El dolor representa, ante todo, un mecanismo protector del cuerpo, porque no es una sensibilidad pura, sino más bien la respuesta al daño tisular que se genera dentro del sistema nervioso. El dolor rápido se percibe 0,1 s después del estímulo, y el lento, 1 s o más después. El dolor lento suele asociarse con un daño tisular y puede denominarse como urente, sordo o crónico. Todos los receptores para el dolor son terminaciones nerviosas libres, cuyo número y densidad alcanza el máximo en la piel, el periostio, las paredes articulares, las superficies articulares, la duramadre y sus reflexiones dentro de la bóveda craneal .
• Los estímulos mecánicos y térmicos tienden a producir un dolor rápido.
• Los estímulos químicos suelen causar un dolor lento, pero no siempre. Algunas de las sustancias químicas que más veces producen dolor son la bradicinina, la serotonina, la histamina, los iones potasio, los ácidos, la acetilcolina y las enzimas proteolíticas. Por otro lado, las prostaglandinas y la sustancia P potencian la sensibilidad de los receptores para el dolor, pero no los excitan de forma directa.
• Los receptores para el dolor se adaptan de forma muy lenta o nula. A veces, la activación de estos receptores se va paulatinamente elevando conforme prosigue el estímulo doloroso; esto se denomina hiperalgesia.
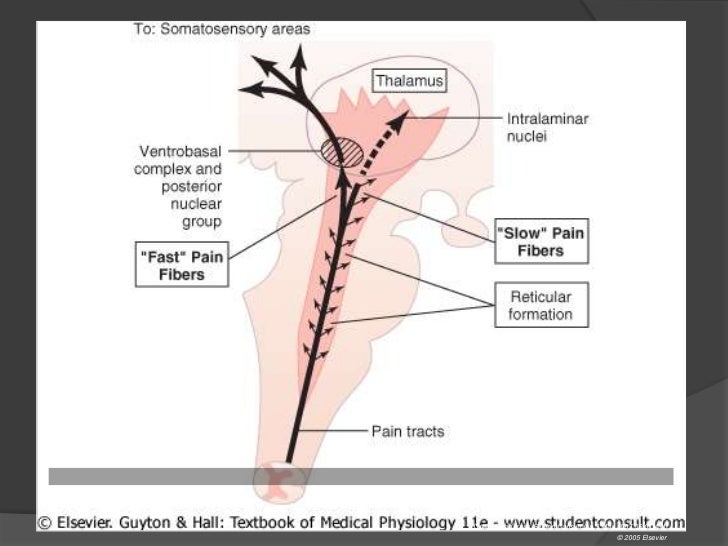
Las señales de dolor rápido desencadenadas por estímulos mecánicos o térmicos son transmitidas por las fibras A6 de los nervios periféricos con velocidades de 6 a 30 mi s. En cambio, las señales lentas de dolor de tipo crónico se vehiculan a través de las fibras de tipo C con velocidades de 0,5 a 2 m/ s. Cuando estos dos tipos de fibras entran en la médula espinal a través de las raíces dorsales, se segregan, de tal suerte que las fibras A8 excitan fundamentalmente las neuronas de la lámina del asta dorsal y las fibras C establecen sinapsis con neuronas de la sustancia gelatinosa. Estas últimas se proyectan en la profundidad de la sustancia gris y activan fundamentalmente neuronas de la lámina V, pero también de las láminas VI y VII. Las neuronas que reciben aferencias de las fibras A8 (dolor rápido) originan el fascículo neoespinotalámico, mientras que las que reciben aferencias de las fibras C crean el fascículo paleoespinotalámico.

El fascículo neoespinotalámico sirve para localizar el dolor. La vía paleoespinotalámica es la más antigua de las dos vías del dolor, en un sentido filogenético y su actividad puede impartir una percepción desagradable del dolor.
La importancia del área somatosensitiva I en la percepción del dolor no está totalmente clara; si se extirpa por completo esta área, no desaparece la percepción del dolor, aunque estas lesiones dificulten la capacidad para interpretar la calidad del dolor y determinar su localización precisa.
Existe una enorme variabilidad en el grado de reacción de las personas a los estímulos dolorosos, en gran medida por el mecanismo que suprime el dolor (analgesia) y que reside en el sistema nervioso central. Este sistema de supresión del dolor consta de tres componentes fundamentales:
• La sustancia gris periacueductal del mesencéfalo y la de la porción rostral de la protuberancia reciben aferencias de las vías ascendentes del dolor, además de proyecciones descendentes desde el hipotálamo y otras regiones del postencéfalo.
• El núcleo magno del rafe (serotonina) y el núcleo paragigantocelular (noradrenalina) del bulbo reciben aferencias de la sustancia gris periacueductal y se proyectan hacia neuronas del asta dorsal de la médula.
• Las interneuronas encefalínicas del asta dorsal reciben aferencias de los axones descendentes serotoninérgicos del núcleo magno del rafe, que entablan contacto sináptico directo con las fibras aferentes del dolor, causando una inhibición presináptica y postsináptica de la señal de entrada. Se cree que este efecto está mediado por el bloqueo de los canales del calcio en la membrana de la terminación de la fibra sensitiva.
Las neuronas de la sustancia gris periacueductal y del núcleo magno del rafe (no las neuronas noradrenérgicas reticulares del bulbo) poseen receptores opioides en la superficie de su membrana que, cuando reciben el estímulo de un opiáceo administrado por vía exógena (analgésico) o de un neurotransmisor opioide endógeno (endorfinas y encefalinas) del cerebro, activan el circuito de supresión del dolor y reducen la percepción de este.
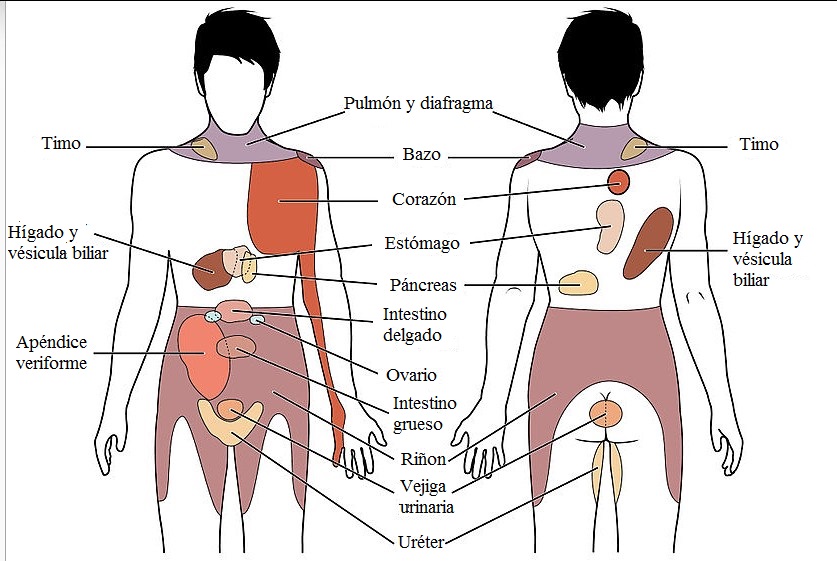
Casi siempre, el dolor referido se debe a señales originadas en un órgano (víscera) o en un tejido internos. El mecanismo no se conoce bien, pero se atribuye a que las fibras de dolor visceral entablan sinapsis con neuronas de la médula que también reciben aferencias de dolor de regiones cutáneas aparentemente no relacionadas con esa estimulación visceral
La hiperalgesia es la acentuación de la sensibilidad a los estímulos dolorosos. El umbral para la activación de los receptores del dolor y la generación subsiguiente de las señales dolorosas disminuye cuando ocurre un daño tisular local o se liberan localmente determinadas sustancias químicas
La infección por virus de un ganglio raquídeo o de un ganglio sensitivo de un nervio craneal puede producir dolor segmentario y un exantema cutáneo intenso en la zona inervada por dicho ganglio, lo que se conoce como herpes zóster (culebrilla).
El encéfalo no es sensible, por sí mismo, al dolor, pero la duramadre y las vainas de los nervios craneales contienen receptores para el dolor que transmiten las señales que circulan por los pares craneales X y XII hasta entrar en los niveles medulares C-2 y C-3. Si se dañan las estructuras sornatosensitivas, el paciente experimenta una sensación de hormigueo o acorchamiento. Las excepciones, son los tics dolorosos y el síndrome de dolor talámico.
La compresión de los senos venosos y el estiramiento de la duramadre o de los vasos sanguíneos y nervios craneales que atraviesan la duramadre produce la cefalea. Cuando se afectan las estructuras situadas por encima de la tienda del cerebelo, el dolor se refiere a la porción central de la cabeza, mientras que la lesión de las estructuras infratentoriales causa cefaleas occipitales. La inflamación meníngea ocasiona, de modo característico, un dolor en toda la cabeza. De la misma manera, si se extrae una pequeña cantidad de líquido cefalorraquídeo (incluso 20 ml) y el paciente no está tumbado, la gravedad hace que el cerebro se «hunda», lo que da lugar a un estiramiento de las meninges, los vasos sanguíneos y los pares craneales, lo que origina una cefalea difusa. Se cree que la cefalea que sigue a una borrachera está causada por la irritación tóxica directa del alcohol sobre las meninges. El estreñimiento también puede producir cefalea por los efectos tóxicos directos de sustancias metabólicas circulantes o por cambios circulatorios asociados a las pérdidas intestinales de líquido.
La tensión emocional puede hacer que los músculos de la cabeza, sobre todo aquellos que se insertan en el cuero cabelludo y en el cuello, se tornen espásticos e irriten los puntos de inserción. La irritación de las estructuras nasales y paranasales de la nariz, extraordinariamente sensibles, explicarían la cefalea sinusal. La dificultad para el enfoque ocular da lugar a una contracción excesiva del músculo ciliar y de los músculos de la cara, en su esfuerzo por agudizar el foco sobre el objeto en cuestión. Se produce así un dolor ocular y facial que suele conocerse como cefalea por esfuerzo ocular.
• Los receptores para el dolor solo se estimulan con el frío o el calor extremos y, en estos casos, se percibe una sensación de dolor y no térmica.
• Los receptores específicos para el calor todavía no se conocen, pero se ha propuesto su existencia en los experimentos psicofísicos; por el momento, se consideran meramente terminaciones nerviosas libres. Las señales de calor son transmitidas por fibras sensitivas de tipo C.
• El receptor para el frío se ha reconocido como una pequeña terminación nerviosa, cuyos extremos sobresalen en la cara basal de las células basales de la epidermis. Las señales de estos receptores son transmitidas por fibras sensitivas de tipo A6. El número de receptores para el frío es de 3 a 10 mayor que los del calor, y su densidad varía entre 15 y 25 por cm2 en los labios, y entre 3 y 5 por cm2 en los dedos de la mano.
Las temperaturas inferiores a 7 ºC y las superiores a 50 ºC activan los receptores para el dolor; estos dos extremos se perciben de manera análoga como un dolor intenso, y no como frío o calor. La temperatura máxima de activación de los receptores para el frío se aproxima a los 24 ºC, y la del calor, a unos 45 ºC. Tanto los receptores para el frío como para el calor se estimulan con temperaturas situadas en el intervalo de 31 a 43 ºC.
los receptores para el frío y el calor responden a la temperatura estacionaria y a los cambios de temperatura. Así se explica por qué una temperatura exterior fría se «percibe» mucho más fría al principio si uno viene de un entorno caliente. El mecanismo estimulador de los receptores térmicos está relacionado, al parecer, con el cambio en el metabolismo de la fibra nerviosa inducido por la variación de temperatura. Se ha comprobado que, con cada variación de la temperatura de 10 ºC, la velocidad de las reacciones químicas intracelulares experimenta un cambio doble.
La densidad de los receptores térmicos en la superficie de la piel es bastante pequeña, por lo que las variaciones de temperatura, si afligen solo a una pequeña región, no se detectan con la misma eficacia que aquellas que actúan sobre una superficie amplia. Si se estimula todo el cuerpo, pueden detectarse cambios térmicos tan pequeños como 0,01 ºC. Las señales térmicas son transmitidas al sistema nervioso central en paralelo con las del dolor.

{kind=link}
{kind=link}
{kind=link}
{kind=link}