La retina se compone de 10 capas o estratos celulares:
- l. Capa pigmentaria
- 2. Capa de conos y bastones
- 3. Membrana limitante externa
- 4. Capa nuclear externa
- 5. Capa plexiforme externa
- 6. Capa nuclear interna
- 7. Capa plexiforme interna
- 8. Capa ganglionar
- 9. Capa de fibras del nervio óptico
- 10. Membrana limitante interna
La fóvea es una región especializada, de aproximadamente 1 mm, situada en el centro de la retina. Dentro del centro de la fóvea existe una zona, denominada fóvea central, con un diámetro de 0,3 mm; esta es la región donde la agudeza visual alcanza el máximo y la capa de fotorreceptores solo contiene conos.

Cada fotorreceptor se compone de:
- 1) un segmento externo
- 2) un segmento interno
- 3) una región nuclear
- 4) el cuerpo o terminal sináptico
Los receptores se denominan bastones o conos, dependiendo, principalmente, de la forma del segmento externo.

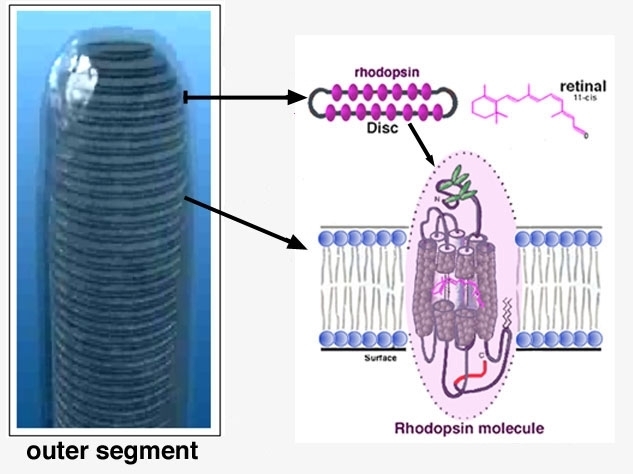
El fotopigmento sensible a la luz rodopsina se localiza en el segmento externo del bastón, mientras que el del cono contiene un material parecido, un pigmento sensible a los colores, la fotopsina. Estos fotopigmentos son proteínas incorporadas a un conjunto apilado de discos membranosos en el segmento externo del receptor, que representa un repliegue de la membrana celular externa.
Los segmentos internos de los bastones y conos son básicamente idénticos y contienen componentes y orgánulos citoplásmicos comunes a otros somas neuronales. Cada núcleo del fotorreceptor se continúa con su propio segmento interno, pero la membrana limitante externa de la retina establece una separación o frontera incompleta entre la capa de los segmentos internos y la capa de los núcleos fotorreceptores (capa nuclear externa).
El cuerpo sináptico contiene elementos como mitocondrias y vesículas sinápticas, habituales en las terminaciones axónicas del cerebro. El pigmento negro melanina de la capa de pigmento reduce la reflexión de la luz por el globo ocular y cumple, por tanto, una función parecida al color negro del interior de los fuelles de una cámara. La importancia de este pigmento se reconoce mejor por su ausencia en las personas albinas. Los albinos casi nunca alcanzan una agudeza visual mayor de 20/100 debido a la enorme reflexión que tiene lugar dentro del globo ocular. La capa de pigmento también almacena grandes cantidades de vitamina A, empleadas en la síntesis de los pigmentos visuales.
La rodopsina se descompone por la energía lumínica. El fotopigmento rodopsina de los bastones se concentra en la porción del segmento externo que sobresale hacia la capa pigmentaria. Esta sustancia es una combinación de la proteína escotopsína y del pigmento carotenoide retinal o, más concretamente, 11-cis retinal. Cuando la rodopsina absorbe la energía lumínica, el retina! se transforma en su configuración todo-trans y comienzan a separarse los componentes retinal y escotopsina. Tras una serie de reacciones extraordinariamente rápidas, el componente retinal se transforma en lumirrodopsina, metarrodopsina I, metarrodopsina II y, por último, escotopsina; y se escinde el todo-trans retinal, Durante este proceso, se cree que la metarrodopsina II se ocupa de la transmisión de los impulsos sucesivos por la retina.
Regeneración de la rodopsina.
En la primera etapa de regeneración de la rodopsina, el todo-trans retinal se convierte en 11-cis retinal, que se combina en seguida con la escotopsina para dar la rodopsina. Existe una segunda vía de formación de la rodopsina basada en la conversión del todo- trans retinal en todo-trans retinol, una forma de vitamina A. El retinol se convierte por vía enzimática en 11-cis retinol y luego en 11-cis retínal, que puede combinarse con la escotopsina para dar rodopsina. Si la retina contiene un exceso de retinal, se transforma en vitamina A, disminuyendo así la cantidad total de rodopsina de la retina.
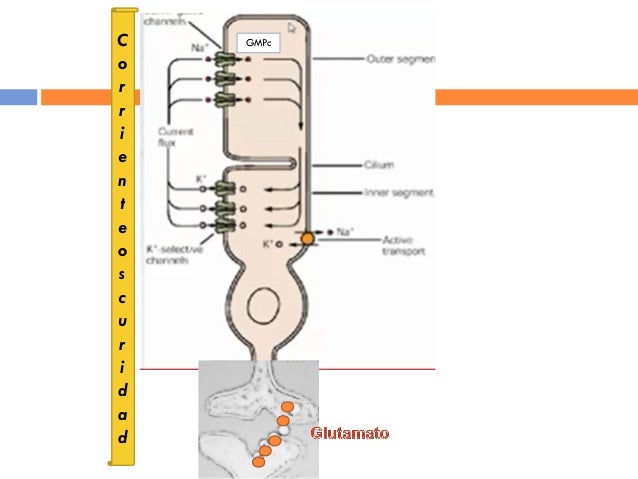
Los bastones siguen un comportamiento muy distinto a otros receptores neurales. En la oscuridad (sin ninguna estimulación lumínica), las membranas del segmento externo de los bastones son «permeables» al sodio, es decir, los iones sodio entran en el segmento externo y cambian el potencial de membrana desde el valor característico de – 70 a -80 m V, hallado en los receptores sensitivos, hasta un valor más positivo de -40 mV. Esta se denomina corriente de sodio o «corriente oscura>> y hace que se libere una pequeña cantidad de transmisor en la oscuridad. Cuando la luz incide en el segmento externo del bastón, las moléculas de rodopsina experimentan la serie de reacciones expuesta con anterioridad, lo que reduce la conductancia del sodio por el segmento externo y también la corriente oscura. Algunos iones sodio siguen siendo bombeados fuera de la membrana celular y la pérdida de estos iones positivos hace que el interior de la célula se torne más negativo; el potencial de membrana se hace más negativo, es decir, se hiperpolariza. Se detiene entonces el flujo del transmisor.
Cuando la luz incide sobre un fotorreceptor, la hiperpolarización pasajera de los bastones alcanza el máximo en unos 0,3 s y dura más de 1 s. Además, la magnitud del potencial de receptor es proporcional al logaritmo de la intensidad lumínica. Este hecho reviste enorme significado funcional, porque permite que el ojo discrimine la intensidad lumínica en un intervalo muchos miles de veces superior al teórico. Todo ello es consecuencia de una cascada química extremadamente sensible que multiplica casi un millón de veces los efectos estimuladores de la siguiente manera. La rodopsina activada (metarrodopsina II) actúa, a modo de enzima, activando numerosas moléculas de transducina, una proteína que se encuentra también en la membrana de los discos del segmento externo. La transducina activada activa, a su vez, la fosfodiesterasa, enzima que hidroliza de inmediato múltiples moléculas de monofosfato de guanosina cíclico (GMPc). Esta pérdida de GMPc hace que se cierren muchos canales de sodio, con lo que el potencial de membrana se va volviendo cada vez más negativo (hiperpolarizado). En el plazo de 1 s, la metarrodopsina ll se inactiva y toda la cascada revierte: el potencial de membrana se torna más despolarizado a medida que se vuelven a abrir los canales de sodio, y el sodio, una vez más, penetra en el segmento externo al restablecerse la corriente oscura. Los conos se comportan de forma parecida, pero el factor de amplificación es entre 30 y 300 veces menor que el de los bastones. La opsina de los conos se denomina fotopsina; y su composición química es distinta a la de la rodopsina, mientras que el componente retinal es exactamente el mismo que el de los bastones. Existen tres tipos de conos, caracterizados cada uno por una fotopsina diferente con una sensibilidad máxima a una longitud de onda determinada de la luz, en la porción azul, verde o roja del espectro lumínico.
Si se efectúa una exposición prolongada a una luz brillante, un porcentaje considerable de las sustancias fotoquímicas de los bastones y de los conos disminuye y gran parte del retinal se transforma en vitamina A. El resultado es un descenso de la sensibilidad global a la luz, que se conoce como adaptación a la luz. Por el contrario, si una persona permanece mucho tiempo en la oscuridad, las opsinas y el retinal se convierten de nuevo en pigmentos fotosensibles. Además, la vitamina A se transforma en retinal, aportando aún más pigmento fotosensible; este proceso se denomina adaptación a la oscuridad y tiene lugar cuatro veces más deprisa en los conos que en los bastones, pese a que los primeros resulten menos sensibles a la oscuridad. Los conos cesan de adaptarse a los pocos minutos, mientras que los bastones, con una adaptación más lenta, continúan haciéndolo durante minutos u horas, y su sensibilidad aumenta dentro de un amplio espectro.
La adaptación puede ocurrir, asimismo, a través de cambios del tamaño pupilar, que puede variar del orden de 30 veces en una fracción de segundo. Los circuitos existentes dentro de la retina y del cerebro también presentan una adaptación neural. Cuando aumenta la intensidad lumínica, también lo hace la transmisión desde la célula bipolar hasta la célula horizontal, la célula amacrina y la célula ganglionar. A pesar de que esta última adaptación es menos potente que los cambios pupilares, la adaptación neural, como la pupilar, ocurre con rapidez.
La utilidad de la adaptación a la luz y a la oscuridad permite que el ojo experimente cambios de sensibilidad por factores de hasta 500.000 a 1 millón, como se reconoce cuando, desde un ambiente muy iluminado, se pasa a una sala oscura. La sensibilidad de la retina es baja, porque está adaptada a la luz y en una habitación oscura se ve poco. Cuando se produce la adaptación a la oscuridad, mejora la visión en esas condiciones. Se cree que la intensidad de la luz solar es 10.000 millones de veces mayor que la intensidad de la luz en una noche estrellada. Sin embargo, el ojo puede operar, hasta cierto punto, en ambas condiciones, debido a su enorme margen de adaptación.
La sensibilidad espectral de los tres tipos de conos depende de las curvas de absorción de la luz de los tres pigmentos respectivos. Todo el color visible (aparte del azul, verde o rojo) es el resultado de la estimulación combinada de dos o más tipos de conos. El sistema nervioso interpreta como color la relación entre la actividad de los tres tipos. Se interpreta como luz blanca una estirnulación equivalente de los conos azules, verdes y rojos.
Si se modifica el color de la luz que ilumina una escena, no se altera sustancialmente la tonalidad del color de esa escena, lo que se conoce como constancia del color, mecanismo atribuido a la corteza visual primaria.
Cuando falta un tipo concreto de cono de la retina, no se logran distinguir ciertos colores. Una persona sin conos rojos sufre protanopia. El espectro global se acorta por el extremo largo de la longitud de onda cuando faltan los conos rojos. La ceguera para los colores rojo y verde es un defecto genético de los hombres transmitido por las mujeres. Los genes del cromosoma X femenino codifican los conos respectivos.
- • Los fotorreceptores se componen de los segmentos externos de los bastones y conos y de los segmentos internos de la capa de fotorreceptores, el cuerpo celular de la capa nuclear externa, y el cuerpo sináptico de la capa plexiforme externa.
- • Las células horizontales, las células bipolares y las células amacrinas reciben aferencias sinápticas de la capa plexiforme externa, poseen somas en la capa nuclear interna y establecen contactos presinápticos con la capa plexiforme interna.
- Las células ganglionares reciben aferencias sinápticas de la capa plexiforme interna, poseen el soma en la capa de células ganglionares y originan axones que viajan dentro de la capa de fibras del nervio óptico.
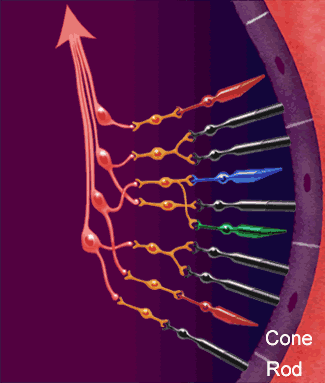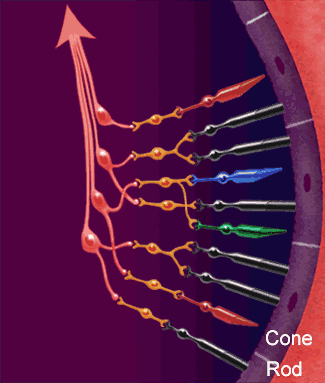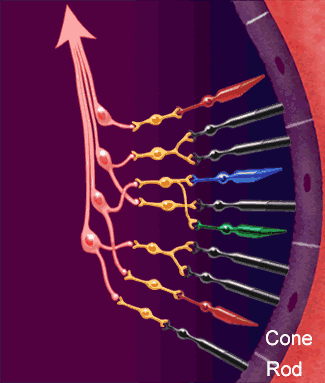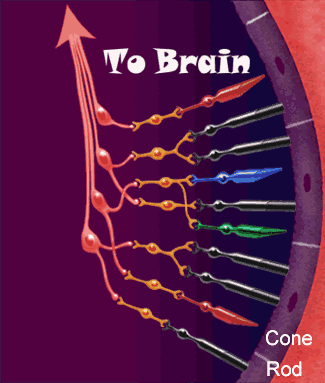
- Las células interplexiformes transmiten las señales en sentido contrario, desde la capa plexiforme interna hasta la externa.
- Las células horizontales pueden intervenir en la capa plexiforme externa, pero las amacrinas ejercen su actividad en la capa plexiforme interna. En las zonas más periféricas de la retina, donde abundan más los bastones, las señales de varios fotorreceptores pueden converger sobre una única neurona bipolar, cuya salida podría dirigirse únicamente a una célula amacrina que se proyecta después hasta una célula ganglionar. Esta constituye la vía de la visión pura con los bastones. Las células horizontales y amacrinas proporcionan una conectividad lateral.
Los neurotransmisores presentes en la retina son:
- el glutamato (utilizado por los bastones y los conos)
- el ácido gama-aminobutírico (GABA)
- la glicina
- la dopamina
- la acetilcolina
- las indolaminas
A partir de los fotorreceptores, las señales se transmiten hasta la capa de células ganglionares exclusivamente por conducción electrotónica (potenciales graduados) y no mediante potenciales de acción. La célula ganglionar es la única neurona de la retina capaz de generar un potencial de acción; así se garantiza que las señales de la retina reflejen exactamente la intensidad lumínica, y ello confiere una mayor flexibilidad a las normas de la retina en su respuesta.
Las prolongaciones de las células horizontales conectan lateralmente con los cuerpos sinápticos de los fotorreceptores y las dendritas de las células bipolares. Los fotorreceptores situados en el centro de un haz de luz reciben la estimulación máxima, mientras que los periféricos son inactivados por las células horizontales, que, a su vez, se activan por el haz luminoso. Se dice que el entorno se inhibe, mientras que la región central se excita ( aunque quizá estos términos no sean precisamente adecuados). Esta es la base de la potenciación del contraste visual. Las células amacrinas también pueden contribuir a potenciar el contraste a través de sus proyecciones laterales hacia la capa plexiforme interna. Curiosamente, mientras que algunas células horizontales poseen axones, las células amacrinas no y, en consecuencia, sus propiedades fisiológicas son extraordinariamente complejas.
Algunas células bipolares se despolarizan cuando la luz estimula sus fotorreceptores o receptores asociados, mientras que otras se hiperpolarizan, dos células bipolares responden simplemente de modo distinto a la liberación de glutamato por el receptor; la primera se excita con el glutamato y la segunda se inhibe. La segunda explicación es que un tipo de célula bipolar recibiría aferencias (excitadoras) directas del fotorreceptor y el otro aferencias inhibidoras indirectas de una célula horizontal. La excitación de algunas células bipolares y la inhibición de otras podrían, asimismo, contribuir al esquema de inhibición lateral.
Se han identificado unas 30 clases de células amacrinas. Algunas responden enérgicamente al comienzo de la estimulación visual, otras al apagado y otras a la iluminación y al apagado. Existe un tipo celular que solo responde a los estímulos en movimiento. Dada la variedad de neurotransmisores empleados por esta categoría de células, no puede efectuarse ninguna generalización en cuanto a su efecto sobre la neurona efectora.
La población de las células ganglionares se divide en las categorías W, X e Y.
- • Las células ganglionares W representan casi el 40% de la totalidad, son pequeñas y el diámetro del soma es de 10 um, la velocidad de transmisión de los potenciales de acción, 8 m/ s, es relativamente lenta. Casi todas sus aferencias provienen de los bastones (a través de las células bipolares y ama crinas) y su campo dendrítico es relativamente amplio. Al parecer, estas células son muy sensibles a los movimientos en el campo visual y, debido a las aferencias predominantes de los bastones, posiblemente se ocupen de la adaptación visual a la oscuridad.
- • Las células ganglionares X son algo más numerosas que las W y representan un 55% de la totalidad. El diámetro del soma varía entre 10 a 15 µm y la velo·cidad de conducción se aproxima a 14 m/s. Estas células poseen campos dendríticos relativamente pequeños, por lo que representan lugares concretos del campo visual. Cada célula X recibe señales de al menos un cono, por lo que esta clase de célula probablemente se ocupe de la visión en color.
- • Las células ganglionares Y son las más grandes, el diámetro del soma llega hasta 35 µm y conducen con una velocidad aproximada de 50 m/s. Como cabe prever, el campo dendrítico es amplio. No obstante, su número es el más bajo, ya que solo representan un 5% de la totalidad. Estas células responden con rapidez a los cambios en cualquier lugar del campo visual (bien en la intensidad o en el movimiento), pero no especifican con exactitud dónde sucede el cambio.
Las fibras del nervio óptico están formadas por axones de las células ganglionares. Aun cuando no sean estimuladas, estas células transmiten potenciales de acción con velocidades que oscilan entre 5 y 40 por segundo. Así pues, las señales visuales se superponen a esta descarga de fondo o espontánea.
Muchas células ganglionares son particularmente sensibles a los cambios en la intensidad de la luz. Algunas responden con una mayor descarga cuando se acentúa la intensidad lumínica y otras incrementan la descarga al reducirse la intensidad de la luz. Estos efectos obedecen a la presencia de células bipolares despolarizantes e hiperpolarizantes. La capacidad de respuesta a la fluctuación en la intensidad lumínica también está muy desarrollada en las regiones periféricas y foveal de la retina.
Se dice que las células ganglionares responden a los límites del contraste más que al valor absoluto de iluminación. Cuando una luz difusa y plana activa los fotorreceptores, las células bipolares despolarizantes proporcionan una salida excitadora, pero las hiperpolarizantes y las células horizontales generan una salida inhibitoria simultánea. Cuando un estímulo luminoso posee un contraste nítido, en el límite entre luz y oscuridad, se hiperpolariza un fotorreceptor de la zona iluminada y se transmite, a través de esa célula bipolar, una señal despolarizante hasta una célula ganglionar que empieza a descargar con más frecuencia. El fotorreceptor vecino de la región oscura se despolariza y se inactiva la línea célula bipolarganglionar. Simultáneamente, una célula horizontal relacionada con el fotorreceptor hiperpolarizado (iluminado) se inactiva porque el fotorreceptor deja de liberar la sustancia transmisora que despolariza la célula horizontal. Así pues, la influencia hiperpolarizante ejercida por esta célula horizontal sobre el fotorreceptor vecino (despolarizado) de la zona oscura desaparece, con lo que el fotorreceptor se despolariza todavía más. La zona oscura se vuelve «más oscura» y la zona de luz se torna «más iluminada» ( es decir, se potencia el contraste).
Algunas células ganglionares son estimuladas por los tres tipos de conos. Se cree que estas células ganglionares emiten una señal luminosa «blanca». Sin embargo, la mayoría de las células ganglionares son estimuladas por la luz de una longitud de onda e inhibidas por otra. Así, la luz roja puede excitar una determinada célula ganglionar, y la verde, inhibirla; este es el mecanismo oponente del color y se atribuye al proceso empleado para discriminar los colores. Como el sustrato de este proceso está presente en la retina, el reconocimiento y la percepción del color pueden comenzar, de hecho, en la retina en el elemento receptor sensitivo primario .

{kind=link}